Muchas de las reacciones catalizadas por las enzimas son muy rápidas debido a que este es el papel más importante que desempeñan, ya que en condiciones fisiológicas normales no se realizarían tan rápido
El catalizador es el encargado de ser quien acelera la llegada al equilibrio de la reacción, estos no cambian la posición del equilibrio de la reacción, mas bien contribruye a que el gasto de energía sea menor
La especificidad es también una propiedad importante y muchas de estas poseen estereoespecificidad ya que solo actúan sobre el estereoisomero del sustrato, atribuyendole la especificidad de reacción que no permite que se creen sustancias tóxicas
Las reacciones enzimáticas funcionan como punto de control en el metabolismo.
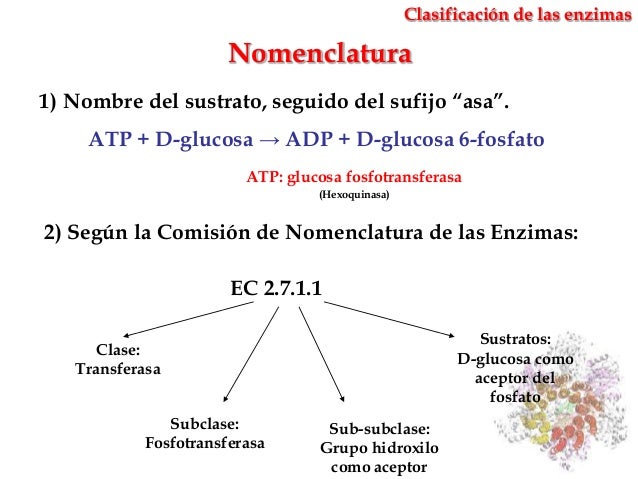
Clasificación

Clasificación
CLASES DE ENZIMAS

UNA PROPIEDAD MUY IMPORTANTE DE LAS ENZIMAS es la cinética
Cinética química: en donde la velocidad dependerá de la concentración del sustrato
Cinética enzimatixa
En primer lugar el sustrato (S) y la enzima (E) se unen y forman un complejo enzima-sustrato (ES). Una vez formado el complejo ES, este o bien se disocia en enzima más el sustrato o se transforma el sustrato en producto formado el complejo enzima-producto (EP), que se disocia para dar enzima (E) más producto (P).
El proceso fue modelizado por Michaelis y Menten y le permitio obtener una ecuación, ecuación de Michaelis-Mente en donde la velocidad de la acción de un enzima es función de la cantidad de sustrato presente, la cantidad de enzima y las características del enzima, la afinidad del enzima por el sustrato y el poder catalítico del enzima:

La Km nos da una idea la afinidad que tiene el enzima por su sustrato, cuanto mayor es Km menor es la afinidad (predominan las formas E y S libres), cuanto menor es Km mayor es la afinidad (predomina la forma ES).

La velocidad máxima Vmáx estima el número de centros activos del enzima. Recordar que hemos definido la Vmáx como la velocidad que obtendríamos cuando todo el enzima se encuentra unido al sustrato. A la constante k2 (poder catalítico del enzima) se la reconoce con el nombre de número de recambio, es el número de moléculas de sustrato convertidas en producto por unidad de tiempo. Por lo que la Vmáx depende de dos cosas, la cantidad de enzima presente y la capacidad catalítica del enzima, es decir la velocidad con que transforma al sustrato.
La representación gráfica de la velocidad en fución de la concentración de sustrato:

Del estudio del modelo se desprende que la velocidad de transformación de sustrato en producto en una reacción enzimática depende:
|  |
Estudia en las siguientes gráficaslos las variaciones de la velocidad en función de los cambios:
Cantidad de sustrato:

Afinidad del enzima por el sustrato y cantidad de enzima o poder catalítico:
Cualquier mecanismo que altere alguno de estos parámetros provocara una modificación de la velocidad enzimática, por lo que una vía podrá ser modulada o controlada. (Ver los diferentes ejemplos de enzimas modulados).
Además no todos los enzimas siguen una cinética de Michales-Menten, algunos presenta una cinetica sigmoidal, los enzimas que presentan modulación alosterica cooperativa, es decir aquellos en el que el sustrato actua como un modulador positivo. Estudia las diferencias de un enzima con modulación alosterica cooperativa positiva y un enzima que sigue la cinética de Michaelis-Menten.
Estudia los efectos en una cinética sigmoidal de los cambios de la Km y la Vmáx:
La velocidad de acción de un enzima también se puede ver afectada por la presencia de inhibidores, que pueden actuar de diferentes maneras. Estudia los efectos sobre la velocidad de la presencia de diferentes inhibidores:

En el caso de que las proteínas diana sean enzimas puede ocurrir dos cosas diferentes: ciertos fármacos actúan como inhibidores de algunas enzimas, es decir la inactivan y no permite que se forme el producto normal o hacen que la catálisis sea mucho más lenta por la misma razón de que el fármaco al ser una sustancia química, se unen al sitio catalítico y no dan cabida al sustrato que normalmente debería unirse. Existen dos tipos de inhibiciones: reversibles que pueden ser competitivas o no competitivas y las inhibiciones irreversibles. En las inhibiciones competitivas tenemos que el fármaco que es introducido a nuestro cuerpo compite contra el sustrato original que es producido en nuestro organismo por el sitio catalítico y dependiendo de cuál de las dos sustancias tenga mayor afinidad por el sitio catalítico se produce una respuesta. Una vez que la respuesta se ha producido, el fármaco deja libre el sitio activo para que la enzima en un futuro pueda unirse a sus sustratos. Por otro lado, aquellas inhibiciones reversibles no competitivas son aquellas en la cual el fármaco ingresa a nuestro organismo y se une a otro sitio de la enzima que no es el activo y cambian su conformación estructural es decir se forma el complejo enzima - inhibidor y también el complejo enzima – inhibidor – sustrato pero como hubo un cambio en la conformación de la enzima, no se producirá una reacción ya que el fármaco unido en otro lado que no es el sitio activo ha dejado sin actividad a la enzima.
En las enzimas alostéricas la unión de un sustrato a un sitio activo puede afectar las propiedades de otros sitios activos en la misma molécula. Un posible resultado de esta interacción entre subunidades es que la unión del sustrato resulte cooperativo, es decir que la unión del sustrato en un sitio activo facilita la unión de los otros sustratos en los sitios activos vecinos.
Además la actividad de una enzima alostérica puede ser alterada por moléculas regulatorias que se unan de manera reversible sitios sobre la proteína que se encuentren en sitios diferentes al sitio activos. Así, las propiedades catalíticas de las enzimas alostéricas pueden ajustarse para cumplir con las demandas inmediatas de la célula. Debido a ello las enzimas alostéricas son los puntos clave de regulación de las vías metabólicas.
Las enzimas reguladas por modificaciones no-covalentes son llamadas de alostéricas. Las mismas son encontradas en casi todas las vías metabólicas y generalmente está catalizando una reacción irreversible localizada en el inicio de la vía. En cuanto a su estructura, son oligoméricas o sea compuestas de varías cadenas polipeptídicas, cada una con una casa de campo activa. La conexión del substrato a la casa de campo activa de una de las subunidades afecta la conformación de las demás, facilitando la conexión de los demás substratos a casas de campo activas.
Las enzimas alostéricas son sensibles reguladores del metabolismo, porque al se conecten la determinados metabolitos celulares su actividad sufre grandes alteraciones. Estos metabolitos también pueden ser llamados de efetuadores o moduladores alostéricos, y pueden ser positivos (aumento de la velocidad de reacción) o negativos (reducción de la velocidad de reacción) de acuerdo con su efecto. Por lo tanto el moduladores alostéricos pueden tutear tanto como inhibidores como activadores de la reacción enzimática.
En la mayoría de las vías metabólicas es común que el producto final tuteé como modulador alostérico negativo de la enzima que cataliza las primeras reacciones de la vía. Por lo tanto cuando la concentración de este producto queda aumentada él va a actuar como un inhibidor alostérico, disminuyendo la velocidad de la vía y su propia producción. Este mecanismo es denominado inhibición por retroalimentación o feedback.
La proteínas con efectos cooperativos tienen curvas de saturación
diferentes a la hipérbola rectangular (la función de saturación de
tipo Michaelis-Menten).
Pueden ser de dos clases:
Efectos cooperativos positivos: Curvas sigmoideas
Efectos cooperativos nega
tivos: Curvas pseudo hiperbólicas
• Efectores Positivos o Activadores. Modifican las constantes
de afinidad (aumentan la afinidad) de tal modo que la unión
de la proteína al ligando tiene lugar a mas bajas
concentraciones del ligando. Los efectores positivos
disminuyen la cooperatividad.
• Efectores Negativos disminuyen las constantes de afinidad.
Aumentan la cooperatividad.
Regulación por modificación covalente
La actividad de una enzima se puede modificar por la fijación covalente y la eliminación de grupos en la cadena polipeptídica.
La regulación por modificación covalente suele ser mas lenta
La modificación covalente de las enzimas reguladas es reversible, pero suele requerirenzimasmodificadorasadicionalesparasuactivacióneinactivación.
Regulación de la piruvato deshidrogenasa de mamíferos. La piruvato deshidrogenasa, es una enzima interconvertible.
• Se inactiva por fosforilacion catalizada por la cinasa piruvatodeshidrogenasa.
• Se reactiva por hidrolisis de su residuo fosfoserina, catalizada por una hidrolasa alostérica llamada fosfatasapiruvatodeshidrogenasa.
Mecanismos de acción
- Sustitución nucleofílica

- Reacciones de ruptura
- Reacciones de oxido- reducción

CATÁLISIS ENZIMÁTICA
COFACOTRES Y VITAMINAS